
Endomycorrhizal diversity associated with some ethnomedicinal plants of Jorhat district, Assam, India
*Corresponding address: liza.handique@yahoo.co.in
Abstract
Endomycorrhizal fungi, particularly arbuscular mycorrhizal fungi (AMF), are highly diverse, forming symbiotic relationships with over 80% of plant species. Predominantly from the Glomeromycota phylum, these fungi include 150- 300+ species within key genera like Glomus, Acaulospora, and Gigaspora, crucial for plant nutrition, soil health, and ecosystem resilience. Their diversity is structural, taxonomic, and functional, with varying colonization abilities and benefits, including improved phosphorus uptake and pathogen resistance. AMF are globally distributed across different ecosystems, with higher richness noted in grasslands and certain medicinal plant areas. In our study, an attempt has been made to study the endomycorrhizal diversity of four ethnomedicinal plants such as Terminalia chebula, Justicia adhatoda, Oroxylum indicum and Melastoma malabathricum. All three plants (Terminalia chebula, Oroxylum indicum and Melastoma malabathricum) had 100% root colonization whereas, Justicia adhatoda had 40 ± 0.67% root colonization. The infection structure study showed that the relative amounts of arbuscular infection were highest in J. adhatoda (100 ± 4.84%), and highest in M. malabathricum for hyphal infection (51 ± 1.33%) and vesicular infection (90 ± 2.53%). The highest mean AMF spore density (198 ± 14.52 spores 50 g soil-1) was obtained from rhizosphere soil of M. malabathricum whereas the lowest spore density (20 ± 1.41 spores 50 g soil-1) was obtained from rhizosphere soil of O. indicum. Increased diversity of arbuscular mycorrhizal fungi (AMF), such as Glomus intraradices, G. mosseae, G. aggregatum, and G. etunicatum, significantly enhances plant community productivity and diversity. This diversity contributes to the resilience and adaptability of host plants in the face of environmental stressors, including drought conditions. Moreover, the inclusion of diverse AMF strains is crucial for promoting plant growth and survival, thereby benefiting sustainable agriculture and ecosystem restoration initiatives. Importantly, belowground mycorrhizal diversity patterns may not always align with those of aboveground plant communities, underscoring the need for conservation efforts focused on underground biodiversity hotspots. The diversity of AMF is influenced by several factors, including the type of host plant and various environmental variables such as soil pH, moisture content, organic matter, and climate conditions.
Keywords: Endomycorrhiza, Glomus, Sustainable agriculture, Soil pH.
Introduction
Endomycorrhiza is a symbiotic association where fungal hyphae (mainly Glomeromycota) penetrate the plant root cortical cells, forming arbuscules and vesicles for nutrient exchange. This widespread relationship improves plant uptake of phosphorus, nitrogen, and water, boosting growth, stress tolerance, and disease resistance. Key aspects of endomycorrhiza include their distinct structure, where the fungi penetrate the root cortical cells of plants, forming vesicles for storage and arbuscules that facilitate nutrient exchange 1. The most prevalent type is vesicular-arbuscular mycorrhizae (VAM), found in over 85% of vascular plant families 2. These fungi are commonly associated with various agricultural crops, such as wheat and rice, as well as vegetables, fruits, and nearly all species of orchids 3. With approximately 120 species (dominated by Glomus, Acaulospora and Scutellospora), endomycorrhizae, especially arbuscular mycorrhizal fungi (AMF), show great diversity in ethnomedicinal plants and promote plant growth and secondary metabolite production. Depending on the pH and moisture content of the soil, these fungi invade plant roots at rates that frequently surpass 70%. Key findings on diversity in ethnomedicinal plants across India indicate that the genus Glomus, including species such as G. fasciculatum and G. mosseae, is the most prevalent among arbuscular mycorrhizal fungi (AMF), followed by Acaulospora and Gigaspora. High colonization rates of AMF, particularly in species like Desmodium pulchellum, contribute significantly to the survival of medicinal plants. Spore density within the rhizosphere exhibits considerable variation, with high levels observed in plants like P. frutescens. The diversity of AMF is affected by ecological factors including soil pH, temperature, and elevation. In the context of ethnomedicine, endomycorrhiza play a crucial role in enhancing the phytochemistry of medicinal plants by increasing the accumulation of bioactive compounds, facilitating nutrient uptake (especially phosphorus), and improving tolerance to environmental stresses such as drought and salinity. Research focusing on regions in India has revealed a diverse range of AMF associated with ethnomedicinal plants, which are used to treat various ailments including diarrhoea, jaundice, and skin infections 4. Mycorrhizal associations are very much beneficial to the plants. They play significant roles in plant establishment in the physical environment. Arbuscular mycorrhizal (AM) fungi in Linum usitatissimum were found to be more stress tolerant. The photosynthetic assimilation of these plants was greater, while the transpiration increase, and root conductance of these plants were decreased than that of nonmycorrhizal plants. Under stress situations, AM fungi assist plants to function normally 5. The large mycelial strands of the fungi of ectomycorrhizae can act as an agent for increasing water movement by binding soil particles together and also encourage water uptake by the plant 6. Recent research has also demonstrated the ability of species of Glomus to transport amino acids (glycine and glutamine) into wheat plants, thereby enhancing nutrient acquisition 7. In some soils with high contents of heavy metals such as zinc (Zn), copper (Cu), manganese (Mn), nickel (Ni) and chromium (Cr), mycorrhizal fungi help plants to grow. Colonisation by these fungi increases the ability of plants to survive in such toxic soils 8, 9, 10. One proposed mechanism of the action of fungi is the sequestration or immobilisation of metals in the fungal hyphae or root tissues thus decreasing the toxicity of the metals in plant shoots 11. Meanwhile, multiple reports indicated that mycorrhizal fungi were also useful in assisting plants to survive in tough soil conditions and under harsh climatic conditions. Haselwandter and Bowen 12 have extensively reviewed this aspect. Terminalia chebula, commonly known as Haritaki, Harad, or Chebulic Myrobalan, is a medium-to-large deciduous tree native to South Asia. Highly revered as the "King of Medicines" in Ayurveda and traditional Tibetan medicine, its dried fruits are extensively used to treat digestive, respiratory, and metabolic disorders. It is also celebrated as one of the three primary ingredients in the famous Ayurvedic rejuvenating formulation, Triphala. Tree characteristics include a height of 25–30 meters with an umbrella-shaped crown and dark brown bark. The oval-shaped leaves are sub-opposite, bright green, and shed in winter. It has small dull white to yellowish-white flowers in spikes that emit a strong odour. The fruit is ellipsoid to ovoid drupes, 1–2 inches long, drying to reveal five longitudinal ridges, ripening from green to yellowish-grey or orange-brown. It serves as a natural laxative, enhancing bowel movements and alleviating chronic constipation. It contains antioxidants that combat aging by neutralizing free radicals and reducing wrinkles. The extracts exhibit strong antibacterial properties against various pathogens and promote gastric health by increasing protective mucus and showing anti-ulcer effects. Additionally, it traditionally helps manage high blood cholesterol and supports blood sugar control in diabetes 13. Justicia adhatoda, commonly known as Malabar Nut, Vasaka, or Adulsa, is a prominent medicinal shrub native to Asia. Belonging to the family Acanthaceae, it grows extensively throughout the seasonally dry tropical biomes stretching from Afghanistan to Indo-China. It is highly revered in traditional medicine systems like Ayurveda, Unani, and Homeopathy for its powerful ability to treat respiratory ailments. An evergreen shrub or small tree, reaching up to 3 meters, features large, lance-shaped green leaves that are bitter in taste. It produces dense, axillary spikes of white flowers with purple or reddish throat markings. The Tamil name, Adathoda, means "untouched by goats," highlighting its resistance to herbivory due to its intense bitterness. The dried leaves of Justicia adhatoda are primarily used as an herbal remedy for respiratory issues. They function as an expectorant and mucolytic, helping to loosen sputum and clear mucus from the respiratory tract. Additionally, they provide bronchial relief by acting as an antispasmodic, aiding in the treatment of chronic cough, bronchitis, asthma, and breathlessness. Notably, the plant contains vasicine, a quinazoline alkaloid that has been utilized in the development of Bromhexine, an over-the-counter mucolytic medication 14. Oroxylum indicum is a night-blooming, deciduous tree native to the tropical and subtropical regions of the Indian subcontinent, China, and Southeast Asia. It belongs to the Bignoniaceae family and is heavily valued in traditional medicine, particularly Ayurveda. The tree is famous for its striking, unusual physical appearance and carries several evocative common names, including the Indian trumpet tree, Midnight Horror, Broken Bones tree, and Tree of Damocles. Growth and stature of the plant range from 10 to 18 metres, characterized by a straight, sparsely branched trunk with rough, greyish-brown bark. It has large compound leaves measuring 0.5 to 1.5 metres long, 2–3 pinnate with broad, ovate leaflets. The reddish-purple or pinkish-yellow trumpet- shaped flowers bloom in large, erect racemes at night, emitting a foul odor to attract nocturnal bats for pollination. The plant produces large, flat, woody seed capsules up to 1 metre long, which crack open to reveal unique, flat seeds with transparent, papery wings. Almost every part of the tree has clinical significance in ethnobotanical practices. In Ayurvedic medicine, the root bark, known as Shyonaka or Sonpatha, is a key ingredient in traditional formulations like Dashamula, an anti-inflammatory tonic, and Chyavanprash. For digestive and liver care, it serves as a bitter tonic for gastric ulcers, diarrhea, dysentery, and jaundice. In respiratory health, practitioners create decoctions from the seeds or bark to alleviate coughs, sore throats, and asthma symptoms. Additionally, a paste made from the bark is applied to joints to relieve pain and swelling from arthritis 15. Melastoma malabathricum, commonly known as the Malabar melastome, Indian rhododendron, or Singapore rhododendron, is a versatile, evergreen tropical shrub belonging to the family Melastomataceae. Despite its common names, it is not a true rhododendron. It is widely celebrated in folklore for its striking purple flowers and extensive medicinal properties, though it is considered a noxious weed in regions like the United States. An erect, branching shrub, 0.5 to 3 metres tall, has distinct four-sided stems with small rigid scales. Its leaves are simple and elliptic-to-lanceolate, featuring 3 to 5 prominent parallel veins, and are rough due to fine, stiff hairs. The shrub produces showy flowers, 5-petaled and measuring 5 to 7.5 cm, in colors ranging from pink to dark purple, with unique dual-colored stamens. A rare white variant named 'Alba' also exists. Its fruit is a capsule that irregularly splits open, revealing dark, sweet, and highly astringent red pulp with tiny U-shaped seeds, which stains the tongue blackish-blue when eaten. In Asian folk medicine, decoctions from the leaves and roots are used to treat various ailments, including diarrhoea, dysentery, stomach aches, and toothaches. Crushed leaves serve as a topical paste for cleaning and healing cuts, wounds, and burns. Additionally, the shrub is recognized for its ability to absorb heavy metals from soil, aiding in environmental cleanup. Its fruits also support wildlife, particularly frugivorous birds like the Double-eyed Fruit-parrot 16. Endomycorrhizal (specifically Arbuscular Mycorrhizal Fungi - AMF) diversity associated with ethnomedicinal plants is critical for enhancing plant growth, nutrient uptake (P, N, Zn, Cu), and the synthesis of bioactive compounds, bridging the gap between agriculture and pharmacology. Studying this diversity helps in identifying beneficial native fungi to improve medicinal plant cultivation, survival in adverse conditions, and conservation of threatened species. Enhanced Secondary Metabolites from AMF colonization positively influence the production of key medicinal compounds in plants. AMF also improve growth and survival by increasing phosphorus uptake, particularly in low-nutrient, high-stress conditions. Identifying native AMF strains can lead to eco-friendly bio-fertilizers, lessening dependency on chemical options for high-value medicinal plants. Additionally, understanding AMF relationships aids in conserving rare medicinal plants and enhances their resistance to soil-borne pathogens.
Materials and method
Study area and collection of plant species
Survey was made at selected areas for the collection of plant specimen. The rhizospheric soil samples of the plant were also collected to investigate and enumerate important microbial associations. The collected plant specimens were preserved in herbarium sheets for further identification. Rhizospheric soil samples (at least three samples at each location) were taken by digging out a small amount of soil (500 g) adjacent to plant roots up to the depth of 15-30 cm. The collected soil samples were kept in pre-sterilized polythene bags at 4 °C to estimate the physico-chemical parameters of soil, mycorrhizal colonization and quantification of VAM fungi as well as actinorhizal and endophytic associations (Figure 1).
Fig 1: Study area
Isolation and quantification of AM spores
AM fungal spores were isolated using the wet sieving and decanting method described by Gerdemann and Nicolson 17, with modifications by Singh and Tiwari 18. 50g soil was added to 500mL water and allowed to mix well. The suspension was then filtered using several sieves of decreasing mesh size. Spores left on the sieves were washed off and transferred to Petri dishes and counted by using a stereo-binocular microscope. The taxonomy keys of Trappe 19, Walker 20, Schenck and Perez 21, Morton and Benny 22 and Mukerji 23 were used to identify the isolated spores. Online resources like databases Mycorrhiza and Glomeromycota were used to provide additional support for identification.
Mycorrhizal quantification
The estimation of spore numbers of VAM was done quantitatively by modified method of Gaur and Adholeya 24. The filter paper with the spores was cut into several pieces and the spores in each piece were counted using a stereo-binocular microscope. The spore population was calculated by adding together the spore counts from each of the sectors.
Colonization of VA Mycorrhizae
The VA mycorrhizae colonization was determined using the quick clearing and staining method of Phillips and Hayman 25. Clean root samples were sliced into 1 cm pieces and kept in 10% potassium hydroxide (KOH) solution for 24 hours at room temperature. The roots were then washed thoroughly with water and the KOH solution was washed off. The root segments were acidified with 1% hydrochloric acid (HCl) for 3-5 minutes then stained with an acid-fast stain – 0.5% trypan blue for 24 hours. Excess dye was removed by staining, with lactophenol. Roots of these plants were stained and placed in the lactic acid:glycerol (1:1) solution and were then examined to determine the extent of mycorrhizal colonization.
Results and discussion
The endomycorrhizal association was studied in four target ethnomedicinal species that is Terminalia chebula, Oroxylum indicum, Melastoma malabathricum and Justicia adhatoda. All three plants (Terminalia chebula, Oroxylum indicum and Melastoma malabathricum) had 100% root colonization whereas, Justicia adhatoda had 40 ± 0.67% root colonization (Table 1). The infection structure study showed that the relative amounts of arbuscular infection were highest in J. adhatoda (100 ± 4.84%), and highest in M. malabathricum for hyphal infection (51 ± 1.33%) and vesicular infection (90 ± 2.53%). The highest mean AMF spore density (198 ± 14.52 spores 50 g soil-1) was obtained from rhizosphere soil of M. malabathricum whereas the lowest spore density (20 ± 1.41 spores 50 g soil-1) was obtained from rhizosphere soil of O. indicum (Table 1). All together 18 arbuscular mycorrhizal fungal taxa were found. The species of Glomus and Acaulospora were the common species of AMF present in the community (Photo plate 1). Acaulospora laevis and Glomus sp.1 was found associated with all four host plant species. Some taxa were host specific, e.g. the Glomalean species Gigaspora gregaria and Glomus constrictum were associated exclusively with M. malabathricum. The proportion of occurrence of these spores in host plants was illustrated graphically as in Fig. 1 (Table 2).
Table 1: The percentage frequency of root colonization, AMF structures, Hyphal, Arbuscular and Vesicular infection, mean spore density and also the type of colonization associated with the target plant species of Jorhat, Assam.
| Sl no | Plant species | Colonization in roots (%) | Hyphal infection (%) | Arbuscular Infection (%) | Vesicular Infection (%) | AMF spore density (per 50 gm of soil) |
|---|---|---|---|---|---|---|
| 1. | Terminalia chebula | 100±0.21 | 22±0.88 | 30±3.45 | 90±1.07 | 154±9.07 |
| 2. | Justicia adhatoda | 40±0.67 | 4.5±1.03 | 100±4.84 | 70±2.08 | 67±3.05 |
| 3. | Oroxylum indicum | 100±0.39 | 46±3.43 | 30±2.36 | 70±1.02 | 20±1.41 |
| 4. | Melastoma malabathricum | 100±0.53 | 51±1.33 | 40±1.77 | 90±2.53 | 198±14.52 |
H=Hyphae; A=Arbuscules; V=Vesicles
± = Standard error

Fig 2: a. Acaulospora laevis, b. Acaulospora lacunosa, c. Acaulospora scrobiculata, d. Funneliformis mosseae, e. Glomus fasciculatum
Table 2: Endomycorrhizal diversity associated with the target plant species of Jorhat, Assam.
| Sl. no. | Spore type | Terminalia chebula | Justicia adhatoda | Oroxylum indicum | Melastoma malabathricum |
|---|---|---|---|---|---|
| 1. | Acaulospora laevis | + | + | + | + |
| 2. | A. pansihalose | - | - | - | + |
| 3. | A. scrobiculata | + | - | - | + |
| 4. | A. birecticulata | - | + | - | + |
| 5. | A. foveata | - | - | + | - |
| 6. | A. lacunosa | + | - | + | - |
| 7. | A. gerdemannii | - | + | - | |
| 8. | Acaulospora sp.1 | + | + | + | + |
| 9. | Gigaspora gregaria | - | - | - | + |
| 10. | Glomus albidum | + | + | - | + |
| 11. | Glomus constrictum | - | - | - | + |
| 12 | Glomus glomerulatum | - | - | - | + |
| 13. | G.mosseae | + | + | - | - |
| 14. | G.clavisporum | - | + | - | - |
| 15 | G. macrocarpon | + | - | - | - |
| 16. | G.pulvinatum | - | + | - | - |
| 17. | Glomus sp.1 | + | + | + | + |
| 18. | Scutellospora sp. | - | + | + | - |
Table 3: Diversity indices of the target plant species.
| Plant species | Shannon-Weiner | Simpson |
|---|---|---|
| Terminalia chebula | 1.953 | 0.840 |
| Justicia adhatoda | 2.212 | 0.882 |
| Oroxylum indicum | 1.373 | 0.650 |
| Melastoma malabathricum | 2.036 | 0.855 |
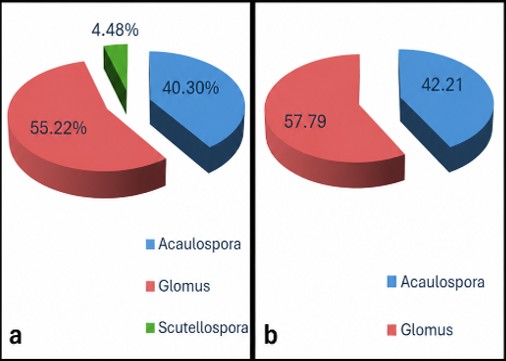
Fig 3 (a)-(d): Percentage occurrence of AM spores in the rhizosphere of the target plant species- (a) Justicia adhatoda (b) Terminalia chebula (c) Oroxylum indicum (d) Melastoma malabathricum.
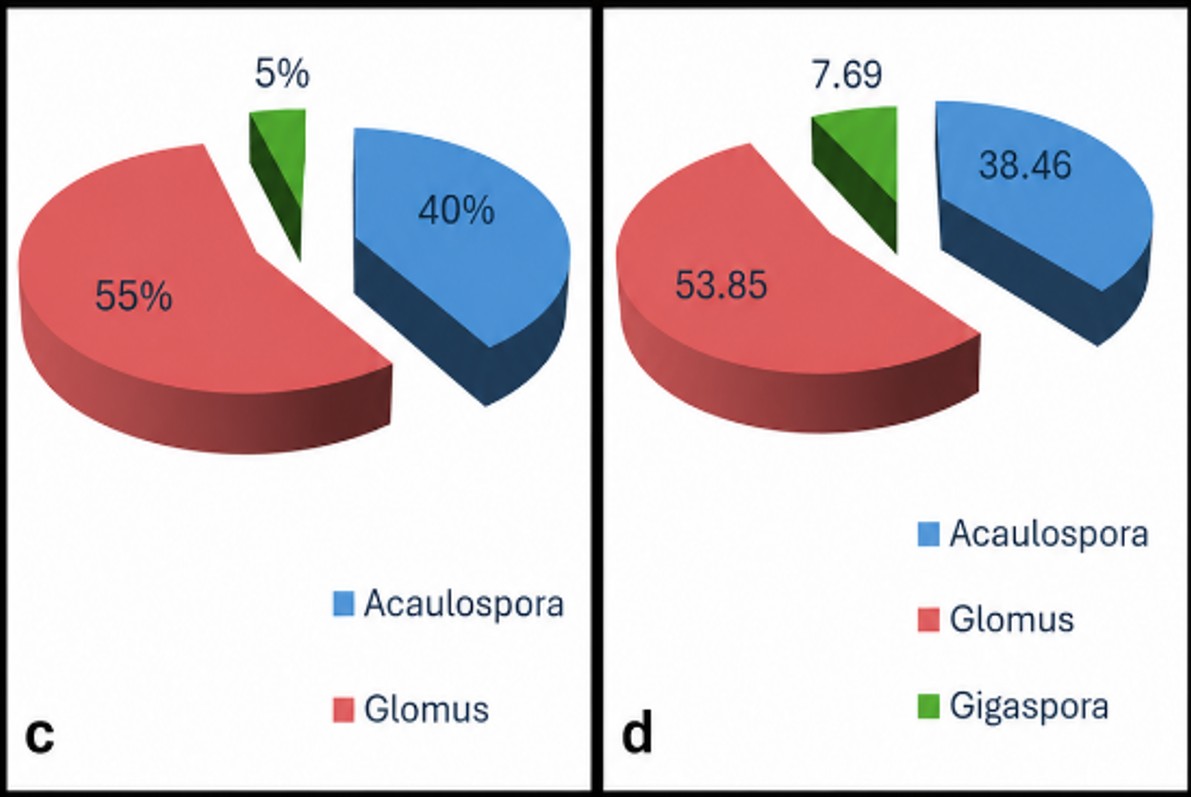
Fig 4: Shannon-Weiner diversity index of the target plant species represented in Radar form.
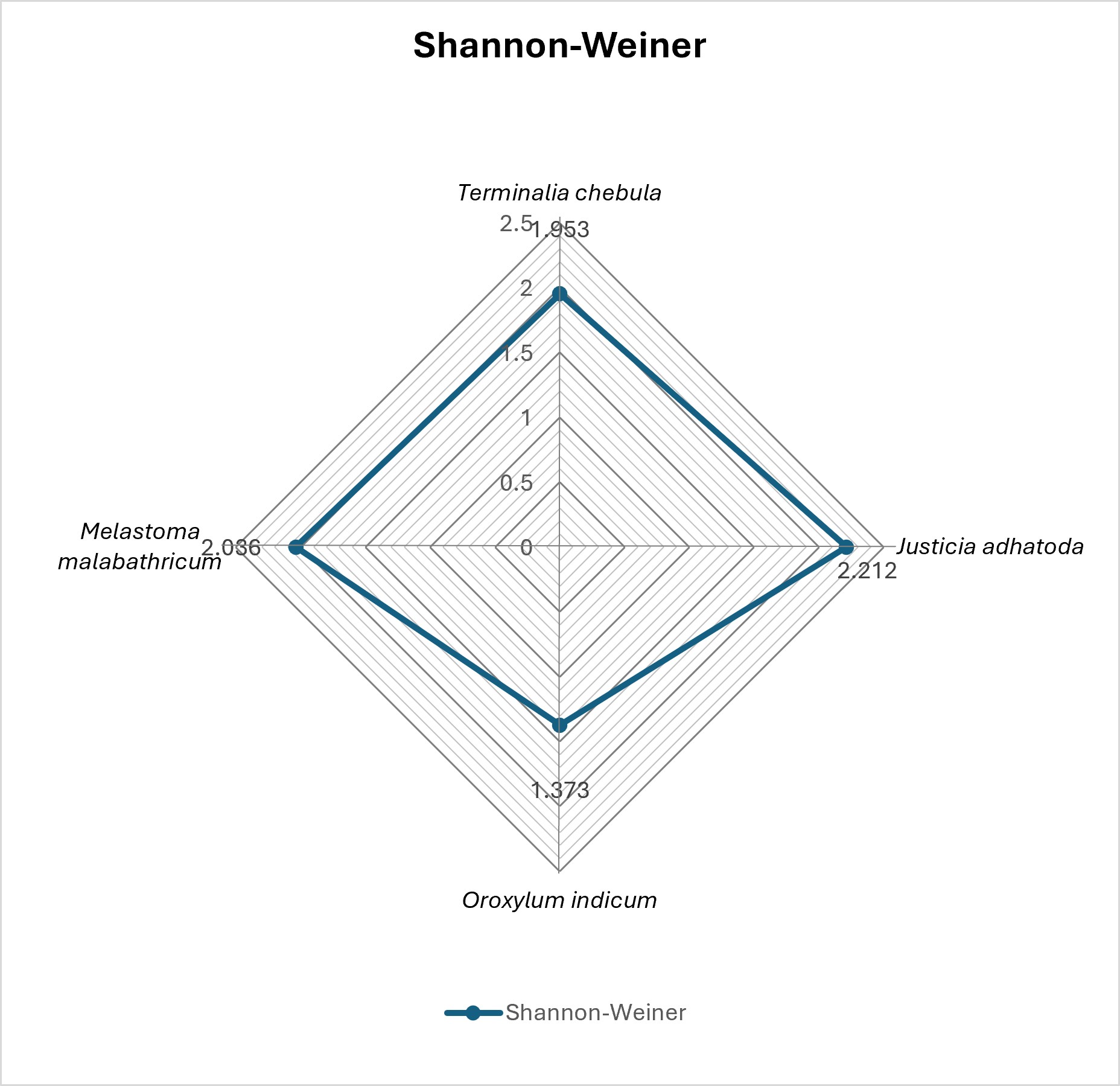
Fig 5: Simpson diversity index of the target plant species represented in Radar form.
Diversity Indices
Community structure was quantified using Shannon–Weiner and Simpson diversity index. The Shannon– Weiner (H′) diversity index for J. adhatoda was the highest and was followed by that of M. malabathricum (2.036). The Simpson-index (D) was the highest for J. adhatoda (0.882) that shows higher AMF diversity and species evenness. This diversity is further highlighted in the representation of the radar shown in Figure 2 and in Figure 3. Arbuscular Mycorrhizal Fungi (AMF) are regarded to be vital as a root symbiotic microbe and participate intensely in nutrient cycling, soil fertility and plant growth in terrestrial ecosystems. The present study showed that occurrence of AMF was found in association with all the selected ethnomedicinal plants of Jorhat, which revealed symbiotic dependency of Jorhat medicinal plants in their rhizosphere. The widespread occurrence of AMF in medicinal plants has been reported earlier for other tropical regions of India 26, 27, 28. AMF are found in various parts of the world and are known to play major physiological and nutritive role in the rhizospheric ecosystem of medicinal plants, which makes them an essential part of the ecosystem. The level of root colonization, structure of infection and the number of spore production were found to vary greatly between species studied. Complete colonization was observed in Terminalia chebula, Oroxylum indicum and Melastoma malabathricum while comparatively lower colonization was observed in Justicia adhatoda. Such differences in colonization could be explained in terms of root morphology and nutrient availability of the host plants and their environment as well as in terms of host-specific rhizospheric interactions. Some other variations were also reported by Garampalli et al. 26 for the case of medicinal plants. Curiously, despite its lower level of root colonization, the J. adhatoda rhizosphere showed high arbuscular infection and higher diversity indices indicating the possibility that more specialized and efficient mycorrhizal association is present in association with J. adhatoda. All the plants studied were in an active mycorrhizal association as evidenced by the presence of hyphae, vesicles and arbuscules. Vesicles are insubstantial energy storage structures, while arbuscules are thought to represent the key point where nutrient exchange occurs between the fungus and host plant. The high level of the arbuscular infection in J. adhatoda can thus be due to active transport of nutrients between fungi and host roots. Likewise the higher prevalence of infection in terms of vesicles and hyphae in M. malabathricum could be related to the fungus establishment and compatibility between the fungus and the host plant flora. There were also significant differences among host plants in AMF spore density. In M. malabathricum, it is assumed that the maximum spore count could be due to optimal rhizospheric conditions, host compatibility and environmental factors affecting spore production while in O. indicum, differences in the environment and production in the host could be the cause of the lower spore count. Some of the edaphic and environmental factors, such as soil pH, soil moisture, phosphorous available, and seasonal fluctuations have been suggested to affect spore abundance in previous studies 26. However, not all the times spore density is related to root colonization as sporulation is related to the physiology of the fungus and environmental conditions 29. There were 18 AMF taxa identified in rhizospheric soils of selected medicinal plants. Most of the species in AMF community were Glomus and Acaulospora. The predominant occurrence of Glomus species seen in this study agrees with previous reports from medicinal plants and tropical soil where Glomus is commonly found as the dominant AMF genus 26, 30. It can be concluded that ecological success of Glomus species may be due to high adaptability, efficient sporulation ability and adaptability to the varying environmental conditions. Acaulospora spp. were also frequently found and may be a characteristic of acidic soil as noted previously by Morton 31. The occurrence of AMF was host-specific, influencing their distribution and diversity, in certain taxa. AMF community composition and species richness has also been shown to be influenced by environmental factors and rhizosphere properties 32. J. adhatoda had relatively higher Shannon–Weiner and Simpson diversity indices showing higher AMF diversity and species evenness in its rhizosphere. The higher the diversity values the more stable the fungal communities and the better the ecological interaction in the soil ecosystem. Climatic factors of the study area might also have played significant role in AMF colonization and diversity. Soil moisture, nutrients and the soil microbial activity are all influenced by temperature and rainfall and hence have a direct impact on the development and sporulation of mycorrhizae. Such a favourable climate during the study period might have favoured the establishment and proliferation of AMF populations in the rhizosphere of the studied medicinal plants. These notes on the effect of the environment on the incidence and diversity of AMF agree with those of Brundrett 33 and Rusan et al. 34. The overall results of the present investigation revealed a diversity of fungal communities of arbuscular mycorrhizae in ethnomedicinal plants occupying the study area. The association of AMF with medicinal plants can play a major role in their nutrient acquisition and adaptive capability in ecosystem and sustainable development. Based on the results obtained in the present study the value of the study may be useful in the further conservation research, production of medicinal plants and creation of ecological-friendly developing strains of bio-fertilizer with the help of native populations of AMF.
References
[1]Brundrett, M. C., & Kendrick, B. (1990). The roots and mycorrhizas of herbaceous woodland plants: II. Structural morphology. New Phytologist, 114(3), 469–479. https://doi.org/10.1111/j.1469-8137.1990.tb00414.x
[2]Tester, M., Smith, S. E., & Smith, F. A. (1987). The phenomenon of nonmycorrhizal plants. Canadian Journal of Botany, 65(3), 419–431. https://doi.org/10.1139/b87-051
[3]Handique, L., & Parkash, V. (2024). Effect of microbial bioinoculation on the bioactive secondary metabolites production of Elaeagnus latifolia L. seedlings. International Journal of Pharmaceutical Sciences and Research, 15(9), 2791–2806. https://doi.org/10.13040/IJPSR.0975-8232.15(9).2791-06
[4]Sharma, K., Singh, M., Srivastava, D. K., & Singh, P. K. (2025). Exploring the diversity, root colonization, and morphology of arbuscular mycorrhizal fungi in Lamiaceae. Journal of Basic Microbiology, 65, e2400379. https://doi.org/10.1002/jobm.202400379
[5]Von Reichenbach, H. G., & Schönbeck, F. (1995). Influence of VA-mycorrhiza on drought tolerance of flax (Linum usitatissimum L.). II: Effect of VA-mycorrhiza on stomatal gas exchange, shoot water potential, phosphorus nutrition and the accumulation of stress metabolites. Angewandte Botanik, 69(5–6), 183–188.
[6]Lamhamedi, M. S., Bernier, P. Y., & André-Fortin, J. (1992). Hydraulic conductance and soil water potential at the soil–root interface of Pinus pinaster seedlings inoculated with different dikaryons of Pisolithus sp. Tree Physiology, 10(3), 231–244.
[7]Hawkins, H. J., Johansen, A., & George, E. (2000). Uptake and transport of organic and inorganic nitrogen by arbuscular mycorrhizal fungi. Plant and Soil, 226(2), 275–285.
[8]Galli, U., Schüepp, H., & Brunold, C. (1994). Heavy metal binding by mycorrhizal fungi. Physiologia Plantarum, 92(2), 364–368.
[9]Hartley, J., Cairney, J. W., & Meharg, A. A. (1997). Do ectomycorrhizal fungi exhibit adaptive tolerance to potentially toxic metals in the environment? Plant and Soil, 189(2), 303–319.
[10]Leyval, C., Turnau, K., & Haselwandter, K. (1997). Effect of heavy metal pollution on mycorrhizal colonization and function: Physiological, ecological and applied aspects. Mycorrhiza, 7(3), 139–153.
[11]Hildebrandt, U., Kaldorf, M., & Bothe, H. (1999). The zinc violet and its colonization by arbuscular mycorrhizal fungi. Journal of Plant Physiology, 154(5–6), 709–717.
[12]Haselwandter, K., & Bowen, G. D. (1996). Mycorrhizal relations in trees for agroforestry and land rehabilitation. Forest Ecology and Management, 81(1–3), 1–17.
[13]Bag, A., Bhattacharyya, S. K., & Chattopadhyay, R. R. (2013). The development of Terminalia chebula Retz. (Combretaceae) in clinical research. Asian Pacific Journal of Tropical Biomedicine, 3(3), 244–252. https://doi.org/10.1016/S2221-1691(13)60059-3
[14]Isha, Kumar, P., & Singh, A. N. (2025). An overview of Justicia adhatoda: A medicinal plant but native invader in India. Conservation, 5(1), Article 2. https://doi.org/10.3390/conservation5010002
[15]Singh, H., & Chaudhary, A. K. (2011). A review on the taxonomy, ethnobotany, chemistry and pharmacology of Oroxylum indicum Vent. Indian Journal of Pharmaceutical Sciences, 73(5), 483–490. https://doi.org/10.4103/0250- 474X.98981
[16]Joffry, S. M., Yob, N. J., Rofiee, M. S., Affandi, M. M., Suhaili, Z., Othman, F., Akim, A. M., Desa, M. N., & Zakaria, Z. A. (2012). Melastoma malabathricum (L.) Smith ethnomedicinal uses, chemical constituents, and pharmacological properties: A review. Evidence-Based Complementary and Alternative Medicine, 2012, Article 258434. https://doi.org/10.1155/2012/258434
[17]Gerdemann, J. W., & Nicolson, T. H. (1963). Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Transactions of the British Mycological Society, 46(2), 235–244. https://doi.org/10.1016/S0007-1536(63)80079-0
[18]Singh, S. S., & Tiwari, S. C. (2001). Modified wet sieving and decanting technique for enhanced recovery of spores of vesicular arbuscular mycorrhizal (VAM) fungi in forest soils. Mycorrhiza News, 12, 12–13.
[19]Trappe, J. M. (1982). Synoptic key to the genera and species of zygomycetous mycorrhizal fungi. Phytopathology, 72, 1107–1108.
[20]Walker, C. (1992). Systematics and taxonomy of arbuscular endomycorrhizal fungi (Glomales): A possible way forward. Agronomie, 12, 887–897.
[21]Schenck, N. C., & Perez, Y. (1990). Manual for the identification of VA mycorrhizal (VAM) fungi. Synergistic Publications.
[22]Morton, J. B., & Benny, G. L. (1990). Revised classification of arbuscular mycorrhizal fungi (Zygomycetes): New order, Glomales; two new suborders, Glomineae and Gigasporineae; and two new families, Acaulosporaceae and Gigasporaceae, with emendation of Glomaceae. Mycotaxon, 37, 471–491.
[23]Mukerji, K. G. (1996). Taxonomy of endomycorrhizal fungi. In K. G. Mukerji, B. Mathur, B. P. Chamola, & P. Chitralekha (Eds.), Advances in Botany (pp. 211–221). APH Publishing Corporation.
[24]Adholeya, A., & Gaur, A. (1994). Estimation of VAM fungal spores in soil. Mycorrhiza News, 6(1), 10–11.
[25]Phillips, J. M., & Hayman, D. S. (1970). Improved procedures for clearing roots and staining parasitic and VAM fungi for rapid assessment of infection. Transactions of the British Mycological Society, 55, 158–161.
[26]Garampalli, R. H., Seema, S., & Sunil, P. (2012). Diversity of arbuscular mycorrhizal fungi associated with some medicinal plants in Western Ghats of Karnataka region, India. World Journal of Science and Technology, 2(1), 13– 20.
[27]Johnson, M. P., & Stephan, R. (2016). Association of arbuscular mycorrhizal fungi and other rhizosphere microbes with different medicinal plants in the calcareous soil of Ariyalur District, India. International Journal of Current Microbiology and Applied Sciences, 5(9), 659–666. https://doi.org/10.20546/ijcmas.2016.509.076
[28]Kumar, A., Mangla, C., Aggarwal, A., & Parkash, V. (2010). Arbuscular mycorrhizal fungal dynamics in the rhizospheric soil of five medicinal plant species. Middle-East Journal of Scientific Research, 6(3), 281–288.
[29]Kalita, R. K., Bora, D. P., & Dutta, D. (2002). Vesicular arbuscular mycorrhizal association with some native plants. Indian Journal of Forestry, 25(2), 143–146.
[30]Selvaraj, T., Murugan, R., & Bhaskaran, C. (2001). Arbuscular mycorrhizal association of Kashini (Cichorium intybus L.) in relation to physico-chemical characters. Mycorrhiza News, 13(2), 14–16.
[31]Morton, J. B. (1986). Three new species of Acaulospora (Endogonaceae) from high aluminum, low pH soils in West Virginia. Mycologia, 78(4), 641–648. https://doi.org/10.1080/00275514.1986.12025298
[32]Abbott, L. K., & Robson, A. D. (1991). Factors influencing the occurrence of vesicular-arbuscular mycorrhizas. Agriculture, Ecosystems & Environment, 35(2–3), 121–150. https://doi.org/10.1016/0167-8809(91)90048-3
[33]Brundrett, M. C. (2002). Co-evolution of roots and mycorrhizas of land plants. New Phytologist, 154(2), 275–304. https://doi.org/10.1046/j.1469-8137.2002.00397.x
[34]Rusan, M. J., Pan, W. L., & Kennedy, A. C. (1998). Seasonal mycorrhizal colonization of winter wheat and its effect on wheat growth under dryland field conditions. Mycorrhiza, 8(3), 139–144. https://doi.org/10.1007/s005720050226